近日,深圳大学化学与环境工程学院周礼杰团队在环境工程领域顶级期刊《Water Research》(Nature Index期刊;影响因子11.4;中科院JCR 1区,TOP期刊)上发表题为《Non-electroactive bacteria behave variously in AnMBR biofilm control using electric field》研究论文。该团队周礼杰副教授为论文第一作者及独立通讯作者,深圳大学为第一作者单位及通讯单位。

电场已经广泛应用于厌氧膜生物反应器。电场的另一个突出应用是直接为细菌的分解代谢过程提供电子,如微生物电解细胞(MECs)。这些电活性细菌可以直接从固体电极获取电子,并通过导电的附体,如纳米线和菌毛,在彼此之间传递电子。因此,电活性细菌通常在不需要任何电场的情况下使用外部电场来富集可溶性电子供体。与电活性细菌占据大量焦点相比,非电活性细菌由于无法直接将电子从电极转移到电极或从电极转移到电极而很少被电化学技术所利用。尽管存在这种限制,但最近的证据表明,在细菌界占主导地位的非电活性细菌可能在电场介导的过程中发挥一些重要作用。例如,低压电场生物反应器中主要含有非电活性细菌中,生物膜内的大多数细菌细胞似乎都是非电活性细菌。因此,我们想知道非电活性细菌是否会受到电场的显著影响。在E-AnMBR操作过程中,非电活性细菌在通过电场控制生物膜形成中起什么作用?
在这项研究中,我们研究了E-AnMBR中非电活性细菌的行为,重点研究了它们对生物膜形成的贡献以及它们对使用各种低压电场控制生物膜形成的反应。通过研究非电活性细菌的群落作用和功能基因丰度,我们旨在阐明在生物膜形成过程中电与非电活性细菌之间的相互作用。了解这些动力学将为非电活性细菌在生物膜控制中的作用提供新的见解,拓宽生物电化学应用的范围,超越传统对电活性生物的重视。
首先使用电位在四个E-AnMBR中没有检测到电流,表明水电解没有发生在实验过程中。如前所述,据报道,一些电活性细菌利用细胞色素c进行电子转移成长。然而,我们分析宏基因组组合基因组(MAGs), 发现Thauera, Desulffomicrobium, andPseudomonas,他们携带完整的或部分反硝化基因,但缺乏细胞色素c编码基因,表明这些MAGs不可能是电活性细菌。此外,生物膜样品的红外扫描显示没有特征在1405cm-1处的吸收波长,这已经是用于指示细胞色素c的存在。综合来看,所有这些证据表明铜网的功能阴极电极可能会产生电磁场排斥细菌(和其他自然产生的有机物)。
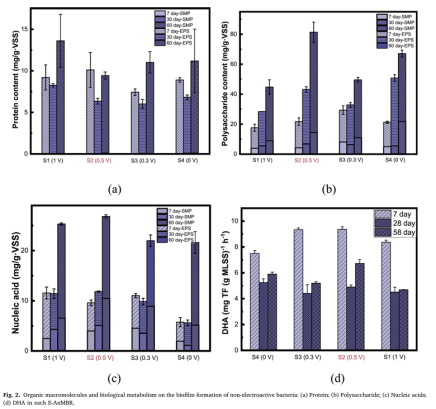
通过SEM观察非电活性菌的生物膜形成情况,生物膜厚度在350 ~ 800 μm之间(图S7)。与S4中的生物膜(468 μm)相比,发现过低(0.3 V)和过高(1 V)的电场电压都阻碍了非电活性细菌在膜表面的生物膜生长。0.5 V电场(S2)显著提高了非电活性细菌的生物膜厚度(780 um),达到S4的1.6倍。此外,S2 (0.5 V)较高的zeta电位绝对值(31.6 mV, p < 0.01)也进一步支持了这一点(图S8)。由此可见,电场显著调节非电活性细菌生物膜的发育,而0.5 V电场则明显刺激非电活性细菌在膜表面的形成。进一步的研究发现,较低的PN/PS被认为增加了非电活性细菌在膜上不可逆的生物膜形成。从图2(a)和(b)可以直观地看出,PN/PS值为S1 (1 V)> S3 (0.3 V)> 54 (0 V)> S2 (0.5 V),S3 (0.3 V)和S1 (1 V)使PN/PS值高于S4,而S2 (0.5 V)使其降低。这一结果与S2的生物膜最厚的观察结果相一致。在0.5 V电场作用下,核酸最高(26.8 mg/g VsS, p < 0.01)。同时,S2的生物膜越厚,其核酸含量也可能越高。这一结果解释了为什么0.5 V电场会加剧由于非电活性细菌生物膜在膜表面形成的不可逆性污染。此外,S2的高脱氢酶活性(DHA)含量(9.36 mg/g VSS, p < 0.01)也支持了这一结论(图d)。在生物处理过程中,高DHA产生更多的代谢废物,从而导致非电活性细菌形成更严重的生物膜。与S4相比,S2的DHA含量增加,而S3和S1的DHA含量则受到抑制。因此,0.5 V电场可以促进非电活性细菌的生物膜形成,并积聚在膜表面。
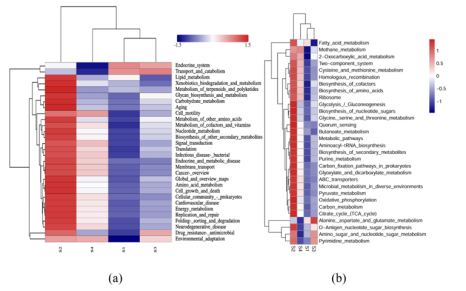
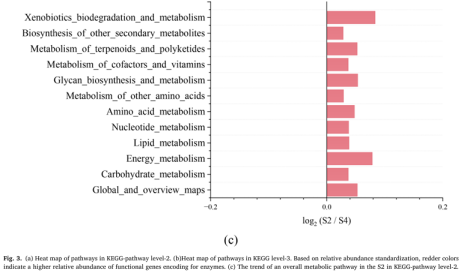
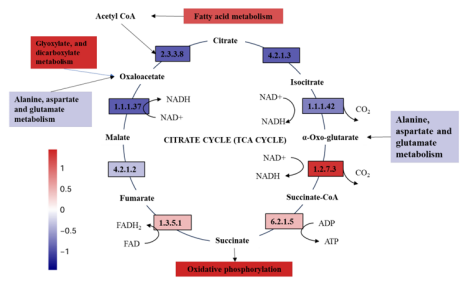
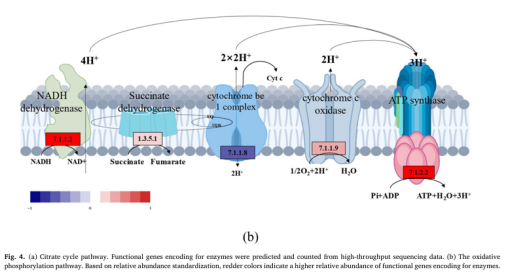
为了理解低电压电场导致E-AnMBR中生物膜变厚这一过程所涉及的机制,我们利用宏基因组技术检测了非电活性细菌群落的功能基因丰度,从而绘制了群落中的元代谢网络图。通过宏基因组分析,从基因水平鉴定了非电活性细菌在电场作用下的代谢途径谱图。功能分析基于KEGG数据库对高丰度代谢途径酶的编码基因进行了标记(图3)。与S4相比,与非电活性细菌生长相关的各种有机物的生物合成和代谢基因(即碳水化合物代谢、脂质代谢和氨基酸代谢)在S2中总体呈上升趋势(图3c),而S1和S3总体呈下降趋势(图3a)。这一结果表明,电场刺激了非电活性细菌的代谢能力。与S4相比,S2的群体感应(QS)基因比SI和S3的要丰富得多(图3b)。QS基因的增加可能有助于细菌之间的信息交换和非电活性细菌的生物膜形成(Sun et al. 2018)。电场还可以干扰微生物群落中的信号通路和通讯机制,包括群体感应,这对生物膜的形成非常重要(Zhou et al. 202b)。氨基酸旺盛的代谢活性(即Glyoxylate和dicarboxylate, Pyruvate,Glycine serine和苏氨酸)进一步证实了这一点(图3a)。基于功能基因预测分析,电场调节碳代谢影响NADH再生和氧化磷酸化编码脂肪酸代谢途径酶的功能基因的增加表明有可能有更多的乙酰辅酶a被燃料注入柠檬酸循环(图4a)。柠檬酸循环途径中编码琥珀酸脱氢酶1.3.5.1、琥珀酸- coa合成酶(6.2.1.5)和2-0xoglutarate reductase[1.2.7.3]的功能基因的高丰度也证实了这一点(图4a)。同时,琥珀酸脱氢酶[1.3.5.1]和2-氧戊二酸还原酶[1.2.7.3]编码的功能基因丰度高,表明会产生更多的氧化磷酸化底物FADH2和NADH。因此,加快了氧化磷酸化的电子传递速度。这表明FADH2和NADH积累,促进了氧化磷酸化过程中的电子转移。事实上,编码氧化磷酸化途径酶的功能基因明显增加(图4b),类似于大多数其他编码参与柠檬酸循环的酶的基因(图4a)。总体而言,电场(0.5 V)通过调节碳代谢影响NADH再生和氧化磷酸化,显著影响非电活性细菌的生物膜形成。3.5. 电场与非电活性细菌代谢途径的相互作用关系代谢途径功能基因、固碳基因、碳代谢基因、TCA循环基因、群体感应基因具有较高的相关系数(图5)。它们的最低相关系数为0.80。说明编码酶的代谢途径基因增加,编码酶的QS基因也会增加。因此,在anmbr中,非电活性细菌是通过电场控制生物膜的关键贡献者。
这项工作得到了广东省自然科学基金-面上项目、广东省普通高校特色创新项目和深圳市高等院校支持计划稳定支持-面上项目的支持。
原文链接:
https://www.sciencedirect.com/science/article/abs/pii/S0043135424015458
撰稿:周礼杰 审核:郭凯、李永亮


 科研动态
科研动态